
Il percorso dove si muovono le bioinformazioni è la matrice extracellulare, MEC, un sistema a tre stadi: stadio solido, stadio fluido e stadio di contatto con le membrane cellulari.
È composta da proteine di adesione, come proteoglicani PG e glicosaminoglicani GAG, proteine strutturali, come fibre elastiche e collagene, e liquido interstiziale (acqua e suoi soluti).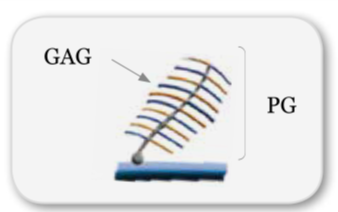
GAG, PG e liquido interstiziale formano un gel dalla consistenza gelatinosa che può essere più o meno solidificabile a seconda delle condizioni fisiologiche e patologiche dell’organismo cui appartiene.
Può infatti stabilizzarsi o passare rapidamente da uno stato di maggiore soluzione (SOL) ad uno di maggiore addensamento (GEL) con spazi più stretti tra le cellule.
I GAG e i PG, per la loro rapida capacità di effettuare cambiamenti strutturali, grazie alla loro parte saccaridica di differenziarsi, di legarsi all’acqua e scambiare ioni, rappresentano i supporti perfetti per l’eliminazione dei fotoni non coerenti e la diffusione di quelli coerenti.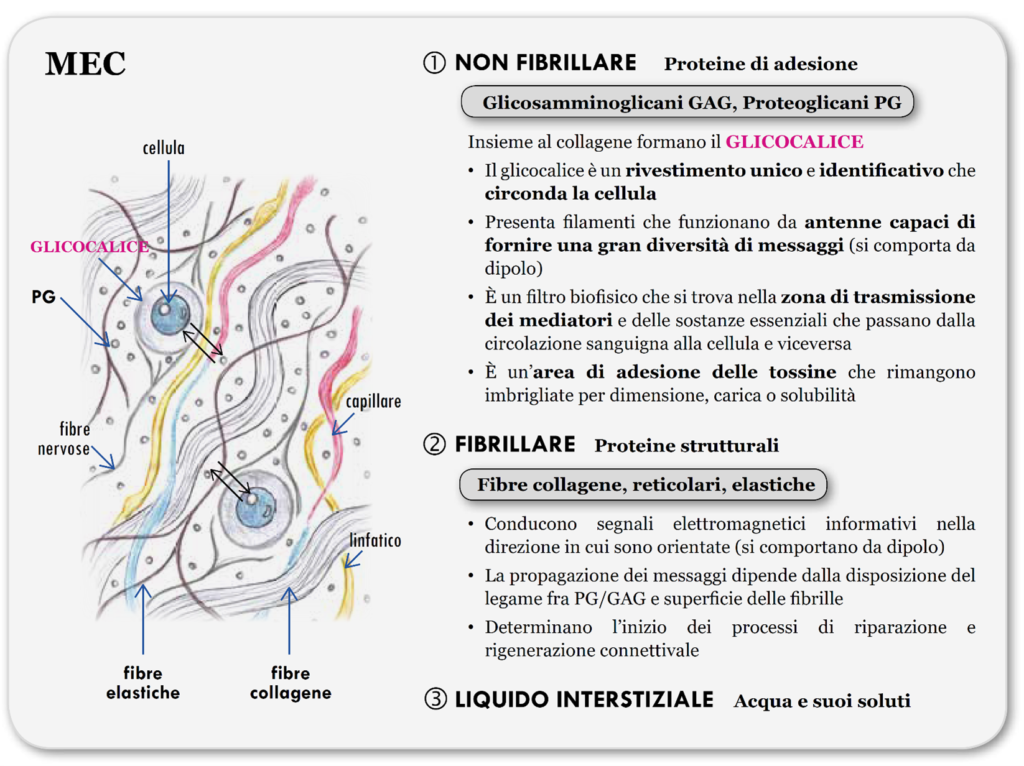
La MEC comprende anche il liquido interstiziale costituito da ACQUA e suoi soluti.
La struttura dell’acqua può essere influenzata nei suoi legami dalle molecole in soluzione che ne modificano il comportamento mostrando configurazioni stechiometriche nuove anche a distanza dal soluto.
In questo modo le molecole d’acqua sono in continuo cambiamento di stato e, quindi, in continuo movimento vibratorio che la rendono capace di attrarre, produrre e riprodurre biomessaggi sia propri che provenienti da altre strutture. Quindi la struttura dell’acqua può essere modificata anche da una molecola disciolta in essa, configurandosi in modo nuovo in base alle informazioni con cui è venuta a contatto.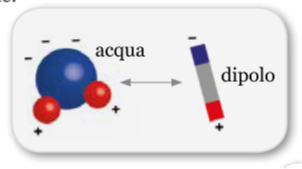
Tali modifiche si mantengono piuttosto stabili se persistono le stesse condizioni che le hanno create. Ogni molecola d’acqua è un dipolo che cede protoni ad altre due molecole d’acqua e ne ricevere da altre due. Essendo i protoni sono divisi fra due atomi di ossigeno, si modificano di continuo variando i loro rapporti fra energia libera e energia nei legami con produzione di un incessante movimento vibratorio.
A livello biologico le molecole di acqua possono comportarsi anche da isolanti e, cioè, collegarsi in catene tramite legami idrogeno, con ossigeno libero, e formare, così, una strada attraverso cui protoni H+ possono saltare da un atomo di ossigeno ad un altro (jumping protonico) coprendo distanze significative all’interno o attorno alle strutture macromolecolari, senza perdita di calore. In pratica, l’acqua biologica fornisce un «rivestimento dinamico» che realizza una minore dispersione della informazione.
L’acqua inoltre può strutturarsi in clusters, o clatrati, grappoli di molecole cristalliformi pentagonali o esagonali che si uniscono e si organizzano come una rete circondando cavità vuote o contenenti macromolecole, frammenti di macromolecole o molecole. La regolare organizzazione dei clusters si deve alla sequenziale disposizione dei legami idrogeno.
I clatrati sono in grado di produrre oscillazioni coerenti in risonanza con i campi magnetici delle strutture chimiche che racchiudono e poi di diffondere all’esterno i biomessaggi che provengono da queste. L’acqua biologica interstiziale conduce quindi la bioinformazione che sembra continuare ad essere inviata anche quando la molecola che la genera non c’è più.
La bioinformazione endogena coerente proveniente dal DNA endonucleare in fase di srotolamento è frutto del corredo cromosomico, è l’espressione fisica di ogni sistema biologico ed è in grado di attivare in modo ordinato e altamente specializzato qualsiasi procedura chimica e fisica e di confrontarsi con qualsiasi informazione esogena.
Questi biomessaggi, disposti in fasci di fotoni coerenti che si susseguono l’uno all’altro, si dirigono in parte verso il nucleo stesso per coordinare la risposta ed in parte entra nel sistema reticolare endoplasmatico, lungo tunnel nel quale il treno fotonico prende velocità, fino ad arrivare alla membrana cellulare che, avendo una struttura isolante, li obbliga a dirigersi verso i glicocalici, vie di uscita verso l’extracellulare.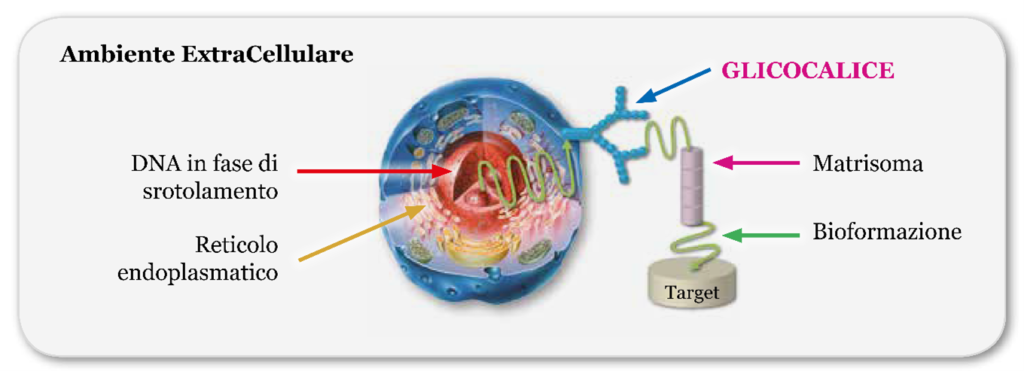
Nella matrice vengono assemblati i matrisomi a seconda delle necessità della bioinformazione e al loro interno i biofotoni viaggiano nel mezzo, la matrice, verso i loro bersagli, posti anche a grande distanza.
La matrice è, quindi, principalmente un grande organo ubiquitario di relazione.
In caso di GEL del sistema, tipico delle flogosi croniche, i messaggi viaggiano meno rapidamente mentre, in caso di SOL del sistema, tipico delle flogosi acute, essendo le cellule più distanti, i fibroblasti ed i macrofagi sono attivati a smantellare le proteine strutturali, le molecole di acqua tendono a muoversi più rapidamente e ad organizzarsi meno in clusters ed i messaggi viaggiano in modo più diretto e rapido.
